Пикорна-, калици-, астро-, тога- и флавивирусы реплицируются наиболее коротким путем: их (+)РНК геном функционирует непосредственно как мРНК.
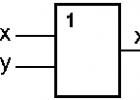
Геномы пикорна- и флавивирусов функционируют как единая полицистрон- ная мРНК, транслирующаяся прямо в единый полипротеин, который впоследствии расщепляется с образованием индивидуальных структурных и неструктурных белков. Одним из них является РНК-зависимая PH К-полимераза, которая реплицирует вирусный геном. Вирусная (+)РНК транскрибируется в комплиментарную (-)РНК-копию, которая служит матрицей для синтеза новых цепей (+)РНК (рис. 6), или может использоваться в качестве мРНК-матриц для синтеза новых минус-цепей и геномной РНК вирусного потомства.
Тога-, корона- и калицивирусы отличаются от пикорнавирусов тем, что на начальном этапе инфекции экспрессируется лишь часть геномной РНК с образованием белков. Последние осуществляют синтез минус-цепи, являющейся матрицей для синтеза различных по размеру классов молекул плюс-РНК. Полипротеины, образующиеся на коротких молекулах мРНК, расщепляются на структурные вирионные белки. Полноразмерные плюс-РНК упаковываются в вирионы.
У тогавирусов транслируется только около 2/3 вирусной РНК (5"-конец); образующийся полипротеин расщепляется на неструктурные белки, которые необходимы для транскрипции и репликации РНК. Вирусная РНК-полимераза синтезирует полноразмерную (-)РНК, на которой затем синтезируются два вида (+)РНК: полноразмерная вирионная РНК, предназначенная для включения в вирионы, и РНК, длина которой равна 1/3, и которая является колинеарной с 3"-концом вирусной РНК и транслируется в полипротеин, который расщепляется на структурные белки (рис. 7). У калицивирусов образуются полигеномные и субгеномные мРНК .
Корона- и артеривирусы демонстрируют необычную стратегию транскрипции: первоначально часть вирионной (+) РНК функционирует как мРНК и транслируется с образованием РНК-полимеразы, которая затем синтезирует полногеномную (-)РНК. На этой (-)РНК транскрибируется гнездо субгеном- ных мРНК с общими 3"-концами. Транслируются только 5"-концевые последовательности каждого члена этого гнезда транскриптов.
Главным отличием вирусов с позитивным геномом является их способность синтезировать ферменты, ответственные за репликацию вирусного генома. Поэтому РНК, выделенная из таких вирусов, инфекционна. Второе отличие состо-

ит в монолитности вирусного генома. Поэтому первичный продукт трансляции обеих РНК (геномной и мРНК) представляет собой единый белок, который в дальнейшем расщепляется на индивидуальные вирусные белки (в том числе структурные) .
Ортомиксо-, бунья- и аренавирусы. Геном представлен (-)РНК. Каждый геномный сегмент транскрибируется отдельно вирионной РНК-транскриптазой и транслируется в один или несколько белков. Эти вирусы характеризуются тем, что их геномная РНК выполняет две матричные функции: в процессе транскрипции и репликации. Транскрипция вирусного генома - первое событие после проникновения вируса в клетку, в результате которого образуются моноцис- тронные мРНК, кодирующие один белок. Репликацию начинают вновь синтезированные вирусные белки, приводящие к образованию плюс-цепи, которая служит матрицей для синтеза геномной (-)РНК (рис. 8).
Следовательно, плюс-транскрипт, функционирующий в качестве мРНК, отличается от (+)РНК, служащей матрицей для вирусного потомства, хотя и первый, и вторая синтезируются на геномной РНК.
Следует отметить, что S-сегмент РНК аренавирусов и некоторых буньявиру- сов является двуполярным, т.е. одна часть сегмента имеет (+) полярность, другая (-) полярность. Стратегия репликации двуполярных РНК-вирусов соответствует полярности их геномов и является смешанной, присущей стратегии репликации (+)РНК и (-)РНК вирусов .
Парамиксо-, рабдо-, фило- и борнавирусы имеют геномную одноцепочечную РНК и РНК-зависимую РНК-полимеразу (транскриптазу), которая транскрибирует пять или более субгеномных (+)РНК, каждая из которых служит в каче-

стве моноцистронной мРНК. В противоположность транскрипции в результате репликации (с помощью той же самой полимеразы, действующей в качестве репликазы) образуются полноразмерные (+)цепи РНК, которые служат матрицей для синтеза новых вирионных (-)цепей РНК. Стратегия репликации борна- вирусов в некоторых деталях является более сложной .
Рео- и бирнавирусы имеют сегментированный двуцепочечный РНК-геном. Негативные цепи каждого сегмента (2-12 сегментов) транскрибируются раздельно в цитоплазме вирионной транскриптазой с образованием мРН К. Эти позитивно полярные (+)РНК служат матрицей для репликации. Образовавшиеся двуспиральные РНК в свою очередь служат матрицей для дальнейшей транскрипции мРНК. Транскрипция генома реовирусов происходит внутри субвирусной частицы с помощью вирионной транскриптазы, причем копируется только одна нить каждого фрагмента двуцепочечной РНК (асимметричная транскрипция). Синтезирующиеся мРНК выходят из сердцевины вириона через 12 полых вершин частично разрушенного капсида. Таким образом, молекулы мРНК реовирусов выполняют две функции. Во-первых, они транслируются, обеспечивая синтез вирусных белков, и, во-вторых, они включаются (по одной молекуле мРНК каждого из 10--12 генов) в состав частиц-предшественниц вирионов. Каждая мРНК служит в этих частицах матрицей для синтеза комплементарной цепи, что ведет к образованию двуцепочечных сегментов вирусного генома (рис. 9).
Молекулы (+)РНК, синтезированные на родительской двуспиральной РНК, связываются с белками и образуют субвирусные частицы, в которых содержится 10-12 фрагментов (+)РНК. Фрагменты (+)РНК в составе субвирусных частиц служат матрицами для синтеза (-)РНК, которые в свободном виде в инфицированных клетках не обнаружены. Полагают, что транскрипция и репликация РНК

реовирусов осуществляется одним ферментом, который модифицируется в процессе репликации вируса .
Ретровирусы. Геном представлен двумя идентичными молекулами (+)РНК. Вместо того чтобы функционировать как мРНК, геном транскрибируется с помощью вирионной обратной транскриптазы, с образованием гибридной молекулы РНК-ДНК. Затем на (-)ДНК синтезируется (+)ДНК и образуется двуспиральная ДНК (другой активностью того же самого фермента) и непрерывно включается в клеточную ДНК. Интегрированная вирусная ДНК (провирус) транскрибируется клеточной РНК-полимеразой II с последующим сплайсингом РНК-транскриптов и расщеплением образующихся белков. Часть вновь синтезированных полноразмерных (+)РНК транскриптов соединяется в пары и образует диплоидные геномы новых вирионов (рис. 10).
Геном ретровирусов состоит из трех генов: (5" - 3") gag, pol и env, которые кодируют, соответственно, внутренние белки вируса, обратную транскриптазу и белки оболочки. Вирус саркомы Рауса имеет в своем геноме четвертый ген - онкоген (src), продукт которого играет ключевую роль в трансформации клеток, но не является необходимым для размножения вируса .
К РНК-содержащим вирусам относятся многие вирусы растений, возбудители заболеваний человека и животных: вирус полиомиелита, вирусы гриппа А, В и С, вирусы паротита (свинки), кори, чумы плотоядных животных (чумки), бешенства, вирус иммунодефицита человека (ВИЧ). В отдельную группу выделяются арбовирусы, которые переносятся членистоногими (клещами, москитами), например, вирусы клещевого энцефалита, желтой лихорадки. Многие РНК-содержащие вирусы вызывают ОРВИ (например, коронавирусы), желудочно-кишечные заболевания (реовирусы птиц, млекопитающих и человека). Некоторые РНК-содержащие вирусы используются в биотехнологии, например, вирусы полиэдроза насекомых.
Вирионы РНК-содержащих вирусов содержат РНК. После проникновения в клетку вирусная РНК становится матрицей для синтеза ДНК и РНК.
Примеры организации генома РНК-содержащих вирусов
- 1. Линейная одноцепочечная мРНК (плюс-цепь) длиной около 4 тн; в виде единой молекулы или в виде нескольких разных молекул. Плюс-цепь сразу же может использоваться для трансляции. Вегетативно-репродуктивная фаза этих вирусов протекает в цитоплазме. В плюс-цепи закодирована РНК-репликаза (РНК-зависимая РНК-полимераза). Представители:
- - Вирус табачной мозаики (ВТМ) - сегментированная РНК. Вирион нитевидный (18х300 нм). ВТМ открыт Д.И. Ивановским в 1982 г.
- - Вирус полиомиелита - несегментированная РНК. Вирионы мелкие, в виде икосаэдра. Капсид белковый.
- - Вирус бешенства. Нитевидный вирион. Имеетсядополнительная липопротеиновая оболочка.
- - Арбовирусы (переносятся членистоногими: клещами, москитами) - вирусы клещевого энцефалита, желтой лихорадки. Морфология и размеры вирионов разнообразны, например, вирус энцефалита содержит 9 белков. Имеется дополнительная липопротеиновая оболочка.
- - Мелкие бактериофаги (с несегментированной РНК).
- 2. Линейная одноцепочечная кРНК (минус-цепь, порядок нуклеотидов комплементарен по отношению к мРНК). Минус-цепь не может служить для трансляции и используется как матрица для синтеза плюс-цепи. Плюс-цепь служит для трансляции вирусных белков и используется как матрица для синтеза вирусной кРНК. Вегетативно-репродуктивная фаза этих вирусов также протекает в цитоплазме.
- - Вирусы гриппа А, В, С. Вирус гриппа А содержит минус-цепь РНК, состоящую из 8 фрагментов. Фрагменты РНК связаны с вирусными белками и образуют спиральный нуклеокапсид. Поверх нуклеокапсида располагается гликолипопротеиновый суперкапсид. В составе вириона 10 белков. В состав суперкапсида входит два белка, определяющих антигенные свойства вируса: гемагглютинин и нейраминидаза. Кроме того, в состав вириона входит уже готовая РНК-репликаза, обеспечивающая синтез плюс-цепи на матрице минус-цепи.
- - Вирусы паротита (свинки), кори, чумы плотоядных животных (чумки). Сферический вирион средних размеров. Имеется дополнительная липопротеиновая оболочка.
- 3. Линейная двухцепочечная РНК
- - Мелкие бактериофаги. Вирионы мелкие, сферические или в виде икосаэдра. Капсид белковый.
- - Вирусы полиэдроза насекомых. Вирионы мелкие, сферические или в виде икосаэдра. Капсид белковый. Используются в биотехнологии (для синтеза интерферона).
- - Реовирусы птиц, млекопитающих и человека. Вирионы мелкие, сферические или в виде икосаэдра. Капсид белковый. Вызывают ОРВИ, желудочно-кишечные заболевания. РНК фрагментированная (10...11 фрагментов), кодирует 11 белков.
- 4. Двелинейные одноцепочечные одинаковые молекулы мРНК длиной около 10 тн. Ретровирусы. Способны интегрироваться в ДНК. В состав вирионов входит фермент обратная транскриптаза (ревертаза). Имеется дополнительная липопротеиновая оболочка. Многие ретровирусы вызывают онкологические заболевания: лейкозы, саркомы, опухоли молочных желез. К ретровирусам относится и вирус иммунодефицита человека, вызывающий СПИД.
- - Вирус иммунодефицита человека (ВИЧ). Содержит одну плюс-цепь РНК, кодирует 13 белков. Сферический вирион. Имеется дополнительная липопротеиновая оболочка, включающая фрагменты мембран человека. Избирательно поражает Т-лимфоциты.
Организация генома митохондрий.
Существуют два типа цитоплазматических ДНК: одни находятся в митохондриях эукариот, другие -в хлоропластах зеленых растений и водорослей. Как и все цитоплазматические элементы, они наследуются по материнской линии, а не по законам Менделя! Большая часть белков этих органелл, закодированная в ядерной ДНК, синтезируется в цитоплазме и затем переходит в органеллу. Однако некоторые белки митохондрий и хлоропластов и все их РНК кодируется в ДНК самих органелл и в них же синтезируются. Таким образом, органеллы - это результат объединенных усилий двух геномов и двух трансляционных аппаратов. РНК-компоненты рибосом органелл, а также тРНК, использующиеся при трансляции, кодируются геномами митохондрий и хлоропластов.
Размеры генома хлоропластов у всех исследованных организмов сходны, тогда как митохондриальные геномы у растений намного больше, чем у животных.
Все митохондрии и хлоропласты содержат по несколько копий собственной геномной ДНК. Эти молекулы ДНК обычно распределены в виде отдельных групп в матриксе митохондрий и в строме хлоропластов, где они прикреплены к внутренней мембране. Способ упаковки ДНК неизвестен. По структуре геном более сходен с бактериальным геномом: например, как и у бактерий, у них нет гистонов.
Геном вирусов включает:
– Структурные гены, которые кодируют белки. Занимают примерно 95 % вирусной хромосомы. Белки вирусов можно разделить на несколько групп: структурные, ферменты, регуляторы.
– Регуляторные последовательности, которые не кодируют белки: промоторы, операторы и терминаторы.
– Прочие некодирующие участки (сайты), в том числе:
– участок attP, обеспечивающий интеграцию вирусной хромосомы в хромосому клетки–хозяина;
– участки cos – липкие концевые участки линейных вирусных хромосом, обеспечивающие замыкание линейной хромосомы в кольцевую форму.
Гены, кодирующие рРНК и тРНК, в геноме вирусов обычно отсутствуют. Однако в геноме крупного фага Т4 имеются гены, кодирующие несколько тРНК.
Геном вирусов отличается высокой плотности упаковки информации. Например, у фага φХ174 в пределах одного гена может располагаться еще один ген. В частности, ген В находится в пределах гена А, а ген Е – в пределах гена D. У мелкого РНК-содержащего фага f2 ген регуляторного белка, блокирующего лизис (созревание вирионов и разрушение клетки), перекрывается с двумя другими генами, удаленными друг от друга.
Особенности вирусов эукариот
У вирусов эукариот обнаружены следующие особенности:
1. Интрон-экзонная структура генов.
2. Модификация белков после синтеза полипротеинов: весь геном транскрибируется в виде одной молекулы мРНК, которая служит матрицей для синтеза полипротеина – одного гигантского инертного белка, и лишь затем происходит расщепление полипротеина на белки, выполняющие определенные функции.
3. Перекрывание генов (обезьяний вирус SV 40, вирус гриппа).
К ДНК-содержащим вирусам относятся многие вирусы бактерий – бактериофаги (или просто фаги). Некоторые мелкие фаги (например, фаг М13) при репродукции не разрушают клетку. Репродукция крупных фагов (например, фага Т–4) приводит к гибели клетки. Фаг Т–4 – это один из наиболее сложно организованных вирусов. Белковый капсид включает не менее 130 белков, образующих головку, воротничок, сократимый хвост, базальную пластинку и хвостовые нити. Такое строение капсида позволяет впрыскивать ДНК в бактериальную клетку через толстую оболочку, поэтому подобные вирусы образно называют «живыми шприцами». Т–фаги могут существовать в виде профага длительное время. К ДНК-содержащим вирусам относятся возбудители многих заболеваний человека и животных: вирусы оспы, герпеса, гепатита В, аденовирусы млекопитающих и человека (вызывают желудочно-кишечные заболевания, ОРВИ, конъюнктивиты), вирусы бородавок человека. К ДНК-содержащим вирусам относятся и некоторые вирусы растений (вирус золотистой мозаики фасоли, вирус мозаики цветной капусты). Некоторые вирусы используются в генной инженерии для переноса генов от одних организмов к другим, например, обезьяний вирус SV 40.
Вирионы ДНК-содержащих вирусов содержат ДНК. Объемом ДНК определяется количество белков в вирионе: один полипептид кодируется отрезком ДНК длиной примерно 1 тысяча нуклеотидов (нуклеотидных пар). После проникновения в клетку вирусная ДНК становится матрицей для синтеза ДНК и РНК.
Примеры организации генома ДНК-содержащих вирусов
1. Кольцевая двухцепочечная ДНК длиной около 5 тпн.
– Обезьяний вирус SV 40. Мелкий эукариотический вирус. Вирионы в виде икосаэдра. Капсид белковый. Используется в генной инженерии как вектор переноса генов. Кодирует 5 белков.
– Вирусы бородавок человека.
2. Кольцевая одноцепочечная ДНК длиной около 5 тн; может быть как кодирующей, так и антикодирующей.
– Мелкие бактериофаги типа М13. Не разрушают клетку. Капсид включает 8 белков.
– Вирус золотистой мозаики фасоли.
3. Линейная двухцепочечная ДНК длиной 30-150 тпн.
– Бактериофаги типа Т4. Вирионы крупные. Белковый капсид из 130 белков включает: головку, хвостовой отдел и хвостовые нити. Эти вирусы могут существовать в виде профага длительное время.
– Аденовирусы млекопитающих и человека. Вирионы средних размеров в виде икосаэдра. Капсиды белковые. Вызывают ОРВИ, конъюнктивиты, желудочно-кишечные заболевания, иногда обладают онкогенными свойствами.
– Вирусы оспы, герпеса и им подобные. Вирионы крупные. Имеется липопротеиновая оболочка.
4. Линейная одноцепочечная ДНК длиной около 5 тн; ДНК может быть как кодирующей, так и антикодирующей. У человека известны как спутники аденовирусов.
5. Двухцепочечная ДНК, замкнутая в кольцо из перекрывающихся сегментов. Длина ДНК – 3-8 тн.
– Вирус гепатита В. Вирион сферический, средних размеров. Имеется дополнительная оболочка из вирусных и клеточных белков. Кодирует 5 белков.
– Вирус мозаики цветной капусты (CaMV). Промотор 35S-RNA (CaMV35S) этого вируса широко используется в традиционной генной инженерии для создания генетических конструкций.
К РНК-содержащим вирусам относятся многие вирусы растений, возбудители заболеваний человека и животных: вирус полиомиелита, вирусы гриппа А, В и С, вирусы паротита (свинки), кори, чумы плотоядных животных (чумки), бешенства, вирус иммунодефицита человека (ВИЧ). В отдельную группу выделяются арбовирусы, которые переносятся членистоногими (клещами, москитами), например, вирусы клещевого энцефалита, желтой лихорадки. Многие РНК-содержащие вирусы вызывают ОРВИ (например, коронавирусы), желудочно-кишечные заболевания (реовирусы птиц, млекопитающих и человека). Некоторые РНК-содержащие вирусы используются в биотехнологии, например, вирусы полиэдроза насекомых.
Вирионы РНК-содержащих вирусов содержат РНК. После проникновения в клетку вирусная РНК становится матрицей для синтеза ДНК и РНК.
Примеры организации генома РНК-содержащих вирусов
1. Линейная одноцепочечная мРНК (плюс–цепь) длиной около 4 тн; в виде единой молекулы или в виде нескольких разных молекул. Плюс-цепь сразу же может использоваться для трансляции. Вегетативно-репродуктивная фаза этих вирусов протекает в цитоплазме. В плюс-цепи закодирована РНК-репликаза (РНК-зависимая РНК-полимераза). Представители:
– Вирус табачной мозаики (ВТМ) – сегментированная РНК. Вирион нитевидный (18х300 нм). ВТМ открыт Д.И. Ивановским в 1982 г.
– Вирус полиомиелита – несегментированная РНК. Вирионы мелкие, в виде икосаэдра. Капсид белковый.
– Вирус бешенства. Нитевидный вирион. Имеется дополнительная липопротеиновая оболочка.
– Арбовирусы (переносятся членистоногими: клещами, москитами) – вирусы клещевого энцефалита, желтой лихорадки. Морфология и размеры вирионов разнообразны, например, вирус энцефалита содержит 9 белков. Имеется дополнительная липопротеиновая оболочка.
– Мелкие бактериофаги (с несегментированной РНК).
2. Линейная одноцепочечная кРНК (минус–цепь, порядок нуклеотидов комплементарен по отношению к мРНК). Минус–цепь не может служить для трансляции и используется как матрица для синтеза плюс–цепи. Плюс-цепь служит для трансляции вирусных белков и используется как матрица для синтеза вирусной кРНК. Вегетативно-репродуктивная фаза этих вирусов также протекает в цитоплазме.
– Вирусы гриппа А, В, С. Вирус гриппа А содержит минус-цепь РНК, состоящую из 8 фрагментов. Фрагменты РНК связаны с вирусными белками и образуют спиральный нуклеокапсид. Поверх нуклеокапсида располагается гликолипопротеиновый суперкапсид. В составе вириона 10 белков. В состав суперкапсида входит два белка, определяющих антигенные свойства вируса: гемагглютинин и нейраминидаза. Кроме того, в состав вириона входит уже готовая РНК-репликаза, обеспечивающая синтез плюс-цепи на матрице минус-цепи.
– Вирусы паротита (свинки), кори, чумы плотоядных животных (чумки). Сферический вирион средних размеров. Имеется дополнительная липопротеиновая оболочка.
3. Линейная двухцепочечная РНК
– Мелкие бактериофаги. Вирионы мелкие, сферические или в виде икосаэдра. Капсид белковый.
– Вирусы полиэдроза насекомых. Вирионы мелкие, сферические или в виде икосаэдра. Капсид белковый. Используются в биотехнологии (для синтеза интерферона).
– Реовирусы птиц, млекопитающих и человека. Вирионы мелкие, сферические или в виде икосаэдра. Капсид белковый. Вызывают ОРВИ, желудочно-кишечные заболевания. РНК фрагментированная (10...11 фрагментов), кодирует 11 белков.
4. Две линейные одноцепочечные одинаковые молекулы мРНК длиной около 10 тн. Ретровирусы. Способны интегрироваться в ДНК. В состав вирионов входит фермент обратная транскриптаза (ревертаза). Имеется дополнительная липопротеиновая оболочка. Многие ретровирусы вызывают онкологические заболевания: лейкозы, саркомы, опухоли молочных желез. К ретровирусам относится и вирус иммунодефицита человека, вызывающий СПИД.
– Вирус иммунодефицита человека (ВИЧ). Содержит одну плюс-цепь РНК, кодирует 13 белков. Сферический вирион. Имеется дополнительная липопротеиновая оболочка, включающая фрагменты мембран человека. Избирательно поражает Т–лимфоциты.
Введение 2 стр.
Глава 1. Строение вирусов. 4 стр.
Глава 2. Разнообразие вирусов. 8 стр.
Глава 3. Биологическая роль вирусов. 11 стр.
Заключение. 14 стр.
Список литературы. 15 стр.
Приложение. 16 стр.
Введение.
"Вирус - это, по существу, часть клетки. Мы считаем вирусами те компоненты клетки, которые достаточно независимы для того, чтобы передаваться другим клеткам, и сравниваем их с другими клеточными компонентами, более прочно связанными со всей системой."
Г. Руска
В 1852 г. русский ботаник Д.И. Ивановский впервые получил инфекционный экстракт из растений табака, пораженных мозаичной болезнью. Когда такой экстракт пропустили через фильтр, способный задерживать бактерии, отфильтрованная жидкость все еще сохраняла инфекционные свойства. В 1898 г. голландец Бейеринк придумал новое слово "вирус" (от латинского слова, означающего "яд"), чтобы обозначить этим термином инфекционную природу некоторых профильтрованных растительных жидкостей. Хотя удалось достичь значительных успехов в получении высокоочищенных проб вирусов и было установлено, что по химической природе это нуклеопротеины (нуклеиновые кислоты, связанные с белками), сами частицы все еще оставались неуловимыми и загадочными, потому что они были слишком малы, чтобы их можно было увидеть с помощью светового микроскопа. Поэтому-то вирусы и оказались в числе первых биологических структур, которые были исследованы в электронном микроскопе сразу же после его изобретения в 30-е годы прошлого столетия.
Вирусы отличаются от микроорганизмов следующими особенностями: 1) они содержат нуклеиновую кислоту только одного типа – или ДНК, или РНК; 2) для их репродукции необходима только нуклеиновая кислота; 3) они не способны размножаться вне живой клетки. Вирусы, таким образом, не являются самостоятельными организмами, а используют для своего размножения живые клетки: их репродукция происходит в клетке-хозяине. Клеточные механизмы нужны как для репликации нуклеиновой кислоты, так и для синтеза белковой оболочки вируса. Развитие вируса приводит к гибели клетки-хозяина. Вне клетки вирус существует в виде вирусной частицы (вириона), которая состоит из нуклеиновой кислоты и белковой оболочки – капсида. Поэтому вирусную частицу называют также нуклеокапсидом. [Шлегель, 1987] В то же время внутриклеточный вирус есть самореплицирующаяся форма, не способная к бинарному делению. Тем самым в определение вируса закладывается принципиальное различие между клеточной формой, воспроизводящейся только из вирусной нуклеиновой кислоты. Однако качественное отличие вирусов от про- и эукариот не ограничивается только одной этой стороной, а включает ряд других: 1) наличие одного типа нуклеиновой кислоты (ДНК или РНК); 2) отсутствие клеточного строения и белоксинтезирующих систем; 3) возможность интеграции в клеточный геном и синхронной с ним репликации.
Вместе с тем вирусы отличаются от обычных репликонов, какими являются молекулы ДНК всех микроорганизмов и любых других клеток, а также плазмид и транспозонов, поскольку упомянутые репликоны являются биомолекулами, которые нельзя отнести к живой материи. [Борисов и др., 1994]
Вирусы распознаются по последствиям своего развития в клетках хозяина. Они разрушают целые комплексы клеток и вызывают поражения тканей, что ведет к появлению некротических пятен или зон лизиса. Обычные хозяева вирусов – это растения, животные и микроорганизмы. [Шлегель, 1987]
Глава 1. Строение вирусов.
Вирусная частица, называемая также вирионом, состоит из нуклеиновой кислоты (ДНК или РНК), окруженной белковой оболочкой. Эту оболочку называют капсидом. Такая единица (капсид + нуклеиновая кислота = нуклеокапсид) может быть «голой», а в других случаях окружена оболочкой. Голым нуклеокапсидами являются, например, частицы вируса табачной мозаики, вируса, вызывающего бородавки, и аденовируса. Дополнительная оболочка окружает вирусы гриппа и герпеса.
Капсид в свою очередь состоит из субъединиц – капсомеров. Он чаще всего имеет симметричное строение. Различают два вида симметрии – спиральную и кубическую. В таблице 1 различные вирусы сгруппированы по их структуре.
Таблица 1. Морфологические классы вирусов.
Спиральная структура
Полиэдрическая структура (икосаэды)
Сложные вирусы (икосаэдрическая головка + спиральный хвост)
Рассмотрим четыре вируса, которые известны как возбудители болезней: два вируса со спиральной симметрией, из них один с голыми частицами (вирус табачной мозаики) и один с дополнительной оболочкой (вирус гриппа), и два типа вирусов с кубической симметрией – с голыми частицами (вирус полиомиелита и другие полиэдрические вирусы) и с оболочкой (вирус герпеса).
Вирус табачной мозаики. Это типичный пример вируса со спиральной симметрией. Его легко выделить из выжатого сока зараженных растений. Частицы представляют собой палочки толщиной 18 нм. Этот палочковидный нуклеокапсид состоит примерно из 2100 капсомеров. Они расположены по винтовой линии и образуют полый цилиндр. Каждый капсомер состоит из одной полипептидной цепи (158 аминокислот, последовательность которых определена). В стенке полого цилиндра между капсомерами помещается цепь РНК, которая тоже идет по винтовой линии.
Вирус гриппа. Частицы вируса гриппа имеют диаметр 110 нм. Нуклеокапсид, как и у вируса табачной мозаики, имеет спиральное строение, но он не палочковидный, а многократно закрученный. Нуклеокапсид окружен оболочкой – фрагментом мембраны клетки-хозяина, из которой вышел вирион. Оболочка имеет на своей наружной стороне шипы, которые служат для адсорбции вириона на поверхности новой клетки-хозяина и содержат мукопротеины и фермент нейраминидиазу. Этот фермент отщепляет от мукопротеинов инфицируемой клетки один компонент – N-ацетилнейраминовую кислоту – и, по-видимому, играет роль в разжижении слизи, покрывающей эпителиальные клетки носоглотки. Размножение вируса происходит внутри клеток. Освобождение вириона напоминает процесс почкования; при этом наружная оболочка вирусной частицы образуется из мембраны клетки-хозяина, которая может быть модифицирована добавлением белков вирусного происхождения (например, нейраминидазы).
Существует много различных разновидностей вируса гриппа. Какую именно ткань будет поражать вирус, зависит от специфичности вируса по отношению к клеткам-хозяевам и от рецепторных свойств клеток. Вирус может вызывать нарушение клеточного метаболизма или даже гибель клетки. Кроме того, он действует как антиген и стимулирует образование антител в организме хозяина. Вирусы, ответственные за большие эпидемии гриппа, отличаются друг от друга по своей вирулентности и патогенности.
Полиэдрические вирусы без наружной оболочки. Многие вирусы, кажущиеся сферическими, на самом деле имеют форму многогранника. Чаще всего это икосаэдр (двадцатигранник) – тело, ограниченное 20 равносторонними треугольниками и имеющее 12 вершин. Капсид икосаэдрического вируса состоит из капсомеров двух типов: в вершинах располагаются пентоны, состоящие из пяти белковых мономеров (протомеров); остальную поверхность граней и ребра образуют гексоны, состоящие из шести протомеров. Построение капсида из капсомеров следует законам кристаллографии; в соответствии с этим наименьший икосаэдрический капсид должен состоять из 12 пентонов, следующий по величине – из 12 пентонов и 20 гексонов. Существуют вирусы из 252 и даже 812 капсомеров.
По принципу икосаэдра построено очень много вирусов: вирусы полиомиелита, ящура, аденовирусы.
Полиэдрические вирусы с наружной оболочкой. Икосаэдр, окруженный оболочкой, - такова форма возбудителей ветряной оспы, опоясывающего лишая и простого герпеса.
Икосаэдрический капсид вируса герпеса состоит из 162 капсомеров. Наружная оболочка, несомненно, образуется из внутренней ядерной мембраны клетки-хозяина. Вирусы герпеса размножаются в ядрах клеток; капсиды новых вирусных частиц одеваются оболочкой из ядерной мембраны, «отпочковываются» от ядра и выводятся наружу по системе эндоплазматического ретикулума.
Ветряная оспа – относительно легкая детская болезнь. Вирус инфицирует верхние дыхательные пути, разносится кровью по всему телу и, закрепляясь в коже, в конечном счете вызывает здесь образование пузырьков. Опоясывающий лишай возникает у частично иммунных лиц; он появляется в результате реактивации вируса ветряной оспы. Таким образом, оба заболевания вызываются одним и тем же вирусом.
Вирус оспы. Вирусы оспы – наиболее крупные из зоопатогенных вирусов. Их частицы устроены совсем не так, как у вирусов четырех представленных выше типов. Они содержат ДНК, белок и несколько липидов, из-за чего их иногда называют комплексными вирионами. Частицы вирусов натуральной оспы и коровьей оспы имеют вид округленных блоков. Они состоят из внутреннего тельца, содержащего двухцепочечную ДНК, двойного слоя, содержащего белок, эллиптических белковых телец и наружной мембраны; частицу обвивают плотно прилегающие к ней нити. Эти вирусные частицы очень устойчивы к высыханию и поэтому чрезвычайно инфекционны. Натуральной оспой могут заболевать только люди и обезьяны. Вирусом коровьей оспы могут заражаться также коровы, кролики и овцы. Оба вируса имеют общие антигены. Поэтому людям профилактически прививают вирус коровьей оспы, который получают от коров и который у человека вызывает весьма слабые симптомы болезни. Такая активная вакцинация приводит к образованию антител, которые обуславливают иммунитет и к натуральной оспе.
Морфология бактериофагов. Строение бактериофагов в основном изучали на примере серии Т Escherichia coli. Колифаг Т 2 состоит из полиэдрической головки длиной 100 нм и отростка, или «хвоста», примерно такой же длины. Поэтому говорят о «составных» вирусах. Головка состоит из капсомеров и содержит внутри ДНК. Количество белка и ДНК примерно одинаково. Отросток фага Т 2 имеет сложное строение. В нем можно различить не менее трех частей: полый стержень, окружающий его сократимый чехол и находящуюся на дистальном конце стержня базальную пластинку с шипами и нитями (от последних зависит специфическая адсорбция на клетке-хозяине). На электронных микрофотографиях, полученных при негативном контрастировании, можно видеть фаговые частицы в двух состояниях: у одних частиц головка очень резко выделяется на электроноплотном фоне и чехол отростка растянут, у других головка мало отличается от фона по плотности и чехол находится в сокращенном состоянии. Первое состояние характерно для активного фага, в головке которого заключена ДНК, второе – для фага, который инъецировал свою ДНК в бактериальную клетку.
Многие бактериофаги имеют более простое строение. В зависимости от формы зрелых фаговых частиц различают ряд типов. Большинство фагов содержит двухцепочечную ДНК. В последние годы, однако, было обнаружено несколько фагов с одноцепочечной ДНК и несколько с одноцепочечной РНК. Содержащие РНК фаги fr, R17, Qβ и другие обладают наименьшими из известных геномов: в них 3500-4500 нуклеотидов. [Шлегель, 1987]
Глава 2. Разнообразие вирусов.
Вирусы составляют царство Vira, которое подразделено по типу нуклеиновой кислоты на два подцарства – рибовирусы и дезоксирибовирусы. Подцарства делятся на семейства, которые в свою очередь подразделяются на роды. Понятие о виде вирусов пока еще четко не сформулировано, так же как и обозначение разных видов.
В качестве таксономических характеристик первостепенное значение придается типу нуклеиновой кислоты и ее молекулярно-биологическим признакам: двунитевая, однонитевая, сегментированная, несегментированная, с повторяющимися и инвертированными последовательностями и др. Однако в практической работе прежде всего используются характеристики вирусов, полученные в результате электронно-микроскопических исследований: морфология, структура и размеры вириона, наличие или отсутствие внешней оболочки (суперкапсида), антигены, внутриядерная или цитоплазматическая локализация и др. Наряду с упомянутыми признаками учитываются и резистентность к температуре, рН, детергентам и т.д.
В настоящее время вирусы человека и животных включены в состав 18 семейств. Принадлежность вирусов к определенным семействам определяется типом нуклеиновой кислоты, структурой, целостностью или фрагментацией генома, а также наличием или отсутствием внешней оболочки. При определении принадлежности к семейству ретровирусов обязательно учитывается наличие обратной транскриптазы. Некоторые таксономические признаки представителей важнейших семейств вирусов человека и животных приведены в таблице 2 (см. приложение 1).
К РНК-содержащим вирусам относится большинство патогенных для человека вирусов. Они отличаются многообразием строения генома, высокими изменчивостью и скоростью эволюции, что приводит к появлению новых возбудителей инфекционных заболеваний. Большинство РНК-геномных вирусов репродуцируется в цитоплазме клетки, хотя некоторые из них на определенных этапах развития локализуются внутри ядра. В настоящее время известно 13 патогенных для человека семейств РНК-геномных вирусов:
1. Семейство Пикорнавирусов.
1.1. Энтеровирусы. Представители: вирусы полиомиелита, Коксаки, ЕСНО, гепатита А и др.
1.2. Риновирусы.
1.3.Афтовирусы. Представители: вирус ящура
2. Семейство Калицивирусов. Представители: вирус Норфолк.
3. Семейство Реовирусов.
3.1. Реовирусы.
3.2. Ротавирусы.
3.3. Орбивирусы.
4. Семейство Ретровирусов.
4.1. Подсемейство Спумавирусов.
4.2. Подсемейство Онковирусов.
4.3. Подсемейство Лентивирусов. Представители: ВИЧ.
5. Семейство Тогавирусов.
5.1. Альфавирусы.
5.2. Вирус краснухи.
6. Семейство Флавивирусов.
6.1. Вирус желтой лихорадки.
6.2. Вирус лихорадки денге.
6.3. Вирус японского энцефалита
6.4. Вирус клещевого энцефалита
6.5. Вирус омской геморрагической лихорадки (ОГЛ).
7. Семейство Буньявирусов.
7.1. Вирус крымской геморрагической лихорадки.
7.2. Вирусы москитных лихорадок.
7.3. Вирус геморрагической лихорадки с почечным синдромом.
8. Семейство Аренавирусов.
8.1. Вирус лимфоцитарного хориоменингита.
8.2. Вирус Ласа.
9. Семейство Филовирусов.
9.1. Вирус Марбург.
9.2. Вирус Эбола.
10. Семейство Рабдовирусов.
10.1. Вирус везикулярного стоматита.
10.2. Вирус бешенства.
11. Семейство Коронавирусов.
12.Семейство Парамиксовирусов.
12.1. Вирусы парагриппа человека (ВПЧГ).
12.2. Вирус паротита.
12.3. Вирус кори.
12.4. Респираторно-синцитиальный (РС) вирус.
13. Семейство Ортомиксовирусов.
13.1. Вирусы гриппа.
Патогенные для человека ДНК-содержащие вирусы входят в состав 6 семейств.
По сравнению с РНК-геномными вирусами они генетически более консервативны, т.е. менее изменчивы, нередко способны к длительной персистенции в организме хозяина. Подавляющее большинство ДНК-содержащих вирусов репродуцируется в ядрах клеток.
1. Семейство Аденовирусов.
2. Семейство Парвовирусов.
3. Семейство Герпесвирусов.
3.1. Альфа-герпесвирусы.
3.2. Бета-герпесвирусы
3.3. Гамма-герпесвирусы
4. Семейство Поксовирусов.
4.1. Вирус натуральной оспы.
4.2. Вирус оспы обезьян.
4.3. Вирус осповакцины (коровьей оспы).
4.4. Вирус контагиозного моллюска.
5.Вирусы гепатита.
5.1. Вирус гепатита А (семейство пикорнавирусов)
5.2. Вирус гепатита В.
5.3. Дельта-вирус.
5.4. Вирус гепатита С.
5.6. Вирус гепатита Е.
6. Онкогенные вирусы.
6.1.Паповавирусы. Представители: Папилломавирусы человека, вирусы полиомы, SV-40.
6.2. Вирусы герпеса.
6.3. Поксивирусы.
6.4. Вирус гепатита В. [Борисов и др., 1994]
Глава 3. Биологическая роль вирусов.
Всем известно, что вирусы играют в основном негативную роль. Но не все знают, что именно вирусы сыграли немаловажную роль в становлении микробиологии и генетики в частности, помогли ученым в изучении свойств и структуры ДНК. Например, при изучении жизненного цикла бактериофагов ученые Лурия и Дельбрюк пришли к предположениям о биологической роли ДНК.
В 1970 г. не известные широкой научной общественности Г.Темин и Д.Балтимор опубликовали в Nature статьи, посвященные обратной транскриптазе (ОТ) – ферменту РНК-содержащих, в том числе раковых, вирусов, которые синтезируют ДНК на матрице РНК, т.е. осуществляют реакцию, обратную той, которую до тех пор наблюдали в клетках.
Открытие обратной транскриптазы позволило выделить первые гены.
Журнал Time назвал Уотсона «охотником за генами». Сам же ученый сказал следующее: «Это захватывающая перспектива. Тридцать лет назад мы не могли и мечтать о том, чтобы узнать структуру генома даже мельчайшего вируса. А сегодня мы уже расшифровали геном вируса СПИДа и почти полностью прочитали геном кишечной палочки объемом в 4,5 млн букв ген-кода. Точное знание детальной структуры генома человека – это восхитительно!».
В нашем геноме много последовательностей, доставшихся нам в «наследство» от ретровирусов. Эти вирусы, к которым относятся вирусы рака и СПИДа, вместо ДНК в качестве наследственного материала содержат РНК. Особенностью ретровирусов является, как уже говорилось, наличие обратной транскриптазы. После синтеза ДНК по РНК вируса вирусный геном встраивается в ДНК хромосом клетки.
Таких ретровирусных последовательностей у нас много. Время от времени они «вырываются» на волю, в результате чего возникает рак (но рак в полном соответствии с законом Менделя проявляется лишь у рецессивных гомозигот, т.е. не более чем в 25% случаев). Совсем недавно было сделано открытие, которое позволяет понять не только механизм встраивания вирусов, но и назначение некодирующих последовательностей ДНК. Оказалось, что для встраивания вируса необходима специфическая последовательность из 14 букв генетического кода. Таким образом, можно надеяться, что вскоре ученые научатся не только блокировать агрессивные ретровирусы, но и целенаправленно «внедрять» нужные гены, и генотерапия из мечты превратится в реальность.
В организме млекопитающих ретровирусы играют и еще одну немаловажную роль. В отношении млекопитающих, у которых плод развивается внутри организма матери, правомерен вопрос: почему иммунная система матери позволяет развиваться организму, который наполовину генетически ей чужероден, поскольку половина генома плода отцовская?
Все дело в ретровирусах, которые блокируют активность иммунных Т-лимфоцитов, ответственных за отторжение органов и тканей, содержащих чужеродные белки, например, после трансплантации органов. Эти ретровирусы активируются в геноме клеток плаценты, которая образуется тканями плода.
Недавно был обнаружен вирус, который блокирует развитие (экспрессию) ретровируса. Если этим вирусом-блокатором заразить беременную мышь, то мышата рождаются нормальными и в срок. Но если его ввести в клетки плаценты, то происходит выкидыш плода, так как активируются Т-лимфоциты матери.
Не стоит забывать, что ретровирусные последовательности возникают также непосредственно на концах хромосом – теломерах. Как известно, теломеры состоят из одноцепочечной ДНК, которая синтезируется ферментом теломеразой по матрице РНК. Считается, что теломеры являются нашими молекулярными часами, поскольку они укорачиваются с каждым клеточным делением. Раньше считалось, что в теломерах нет генов, однако расшифровка генома показала, что генов там довольно много и они активны в детстве и молодом возрасте, постепенно «угасая» по мере старения организма.
Способы передачи вирусных заболеваний.
Капельная инфекция - самый обычный способ распространения респираторных заболеваний. При кашле и чихании в воздух выбрасываются миллионы крошечных капелек жидкости (слизи и слюны). Эти капли вместе с находящимися в них живыми вирусами могут вдохнуть другие люди, особенно в местах скопления большого количества народа, к тому же еще и плохо вентилируемых. Стандартные гигиенические приемы для защиты от капельной инфекции правильное пользование носовыми платками и проветривание комнат.
Некоторые микроорганизмы, такие, как вирус оспы, очень устойчивы к высыханию и сохраняются в пыли, содержащей высохшие остатки капель. Даже при разговоре изо рта вылетают микроскопические брызги слюны, поэтому подобного рода инфекции очень трудно предотвратить, особенно если микроорганизм очень вирулентен (заразен).
Контагиозная передача (при непосредственном физическом контакте). В результате непосредственного физического контакта с больными людьми или животными передаются сравнительно немногие болезни. Сюда прежде всего относятся венерические (т. е. передающиеся половым путем) болезни, такие, как СПИД. К контагиозным вирусным болезням относятся обычные бородавки (папилломавирус) и простой герпес - "лихорадка" на губах.
Переносчик - это любой живой организм, который разносит инфекцию. Он получает инфекционное начало от организма, называемого резервуаром или носителем. Вирус бешенства сохраняется и передается одним и тем же животным, например собакой или летучей мышью. В этих случаях переносчик выступает в качестве второго хозяина, в теле которого может размножаться патогенный микроорганизм. Насекомые могут переносить возбудителей болезней на наружных покровах тела.
Заключение.
Вирусы играют довольно важную роль в биосфере – они выступают как бы одним из факторов естественного отбора, позволяют стабилизировать численность популяций живых организмов. Более сильные организмы вырабатывают антитела и вместе с ними иммунитет, более слабые – погибают. Это позволяет более приспособленным давать потомство с уже сформированным иммунитетом к данному вирусу.
Вирусы помогли ученым изучить роль ДНК в организме. А также было выяснено, что ретровирусы не только являются возбудителями страшных заболеваний: рака и СПИДа, - но и позволяют всем нам появиться на свет, т.к. они блокируют иммунные Т-лимфоциты матери, что не позволяет плоду быть отторгнутым.
Таким образом, вирусы играют не только отрицательную роль, о которой все люди знают не понаслышке, но и исключительно положительную. Дальнейшее исследование этих загадочных «живых – неживых» организмов не только необходимо для человечества, чтобы бороться с неизлечимыми болезнями, вызываемыми вирусами, но и, я думаю, не менее интересно.
Список литературы.
1. Борисов Л.Б., Смирнова А.М., Фрейдлин И.С. и др. «Медицинская микробиология, вирусология, иммунология». – М.: «Медицина», 1994
2. Шлегель Г. «Общая микробиология». – М.: «Мир», 1987
3. www.medicinform.net
4. www. Elite-genetix.ru
Приложение 1.
Таблица 2. Некоторые таксономические признаки представителей важнейших семейств вирусов человека и животных.
| Таксономический признак |
Семейство |
Важнейшие представители |
| I. ДНК-содержащие вирусы |
||
| Двунитевая ДНК |
Аденовирусы |
Аденовирусы |
| Отсутствие внешней оболочки |
Паповавирусы |
Вирусы папилломы, полиомы и бородавок человека |
| Однонитевая ДНК Отсутствие внешней оболочки |
Парвовирусы |
Аденассоциированные вирусы |
| Двунитевая ДНК. Наличие внешней оболочки |
Герпесвирусы |
Вирусы простого герпеса, цитомегалии, ветряной оспы |
| Гепаднавирусы |
Вирус гепатита В |
|
| Поксовирусы |
Вирус натуральной оспы, осповакцины |
|
| II. РНК-содержащие вирусы |
||
| Плюс-однонитевая РНК. Отсутствие внешней оболочки |
Пикорнавирусы |
Вирусы полиомиелита, Коксаки, ЕСНО, вирус гепатита А. |
| Калицивирусы |
Вирусы гастроэнтерита детей (Норфолк) |
|
| Двунитевая РНК. Отсутствие внешней оболочки |
Реовирусы |
Реовирусы, ротавирусы, орбвирусы |
| Наличие обратной транскриптазы |
Ретровирусы |
ВИЧ, вирусы Т-лейкоза, онковирусы |
| Плюс-однонитевая РНК. Наличие внешней оболочки |
Тогавирусы |
Вирусы омской геморрагической лихорадки, краснухи |
| Плюс-нитевая РНК (позитивный геном) |
Флавивирусы |
Вирусы клещевого энцефалита, лихорадка денге, желтой лихорадки. |
| Минус-однонитевая РНК. |
Буньявирусы |
Вирусы Буньямвера, крымской геморрагической лихорадки |
| Аренавирусы |
Вирусы лимфоцитарного хориоменингита, болезни Лассо |
|
| Рабдовирусы |
Вирусы бешенства, везикулярного стоматита |
|
| Двунитевая РНК. Наличие внешней облочки |
Парамиксовирусы |
Вирусы парагриппа, паротита, кори, РСВ |
| Ортомиксовирусы |
Вирусы гриппа человека, животных, птиц |
|
Открытие вирусов , вызывающих злокачественные опухоли у животных, произошло еще на рубеже XIX и XX веков. В 1910 г. Пейтон Рауш обнаружил, что бесклеточный фильтрат из тканей птичьей саркомы может вызывать развитие аналогичной саркомы у цыплят. Примерно в это же время была доказана вирусная природа птичьего миелобластоза. Позднее было обнаружено, что часто проходит весьма значительный латентный период между инфицированием вирусом и развитием рака.
Тем не менее вплоть до 1960-х годов не существовало четких доказательств того, что инкорпорация вирусной ДНК в геном клетки является необходимым условием развития злокачественной трансформации, как не было и случаев выделения вирусной ДНК из раковых клеток.
В настоящее время выделяют два типа вирусных онкогенов . Оба этих типа онкогенов встраиваются в клеточную ДНК. Вирусы первого типа несут онкогены, которые вызывают быструю злокачественную «трансформацию» клеток в культурах in vitro, а в организме вызывают развитие опухолей. При втором типе вирус действует медленнее и для развития опухоли требуется значительное время. Вирусы второго типа не вызывают злокачественной трансформации клеток в культурах in vitro.
РНК-содержащие вирусы вызывают развитие целого ряда различных опухолей у животных, при этом наиболее распространена индукция этими вирусами лимфом, лейкемий и сарком. Типичная структура таких вирусов - это две идентичные цепочки молекул РНК в сочетании с ферментом обратная транскриптаза, одетых в гликопротеиновую оболочку. При инфицировании вирусом его обратная транскриптаза заставляет клетки синтезировать ДНК, комплементарную вирусной РНК.
Эта ДНК затем встраивается в клеточные хромосомы, и на ее основе клетка уже сама начинает синтез новых вирусных белков, вирусных обратных транскриптаз и элементов гликопротеиновой оболочки. Из-за механизма их действия этот тип вирусов получил название ретровирусы. Все они имеют весьма схожий внешний вид на электронных микрофотографиях и являются самыми маленькими из известных вирусов.
Некоторые из ретровирусов (например, вирусы птичьего лейкоза, кошачьей и мышиной лейкемии) содержат в себе только три гена и обладают очень длительным инкубационным периодом с момента заражения до возникновения опухоли. Другие вирусы (например, вирус саркомы Рауса (ВСР)) вызывают очень быструю злокачественную трансформацию и могут быть выделены из культуры опухолевых клеток.
Показано, что вирус ВСР содержит особый ген (v-src), способный вызывать трансформацию фибробластов in vitro. Этот ген кодирует наработку протеинкиназы, которая фосфорилирует тирозин. К сожалению, действие этой протеинкиназы запускает целый каскад различных метаболических процессов, и очень сложно оценить, какой именно из них ведет к злокачественной трансформации.
В настоящее время известно, что и нормальные, и злокачественные клетки содержат в своем генотипе участки ДНК, сходные или идентичные ряду последовательностей онкогенных РНК-содержащих вирусов. Такие участки получили название клеточных прото-онкогенов (чтобы отличать их от вирусных онкогенов). Постулируется, что активация этих участков, возникающая в результате канцерогенного воздействия, запускает целую цепочку событий, ведущую в итоге к злокачественной трансформации клетки. Считается также, что ретровирусы инкорпорировали эти клеточные участки в свой геном в процессе эволюции.
В настоящее время мы лучше понимаем механизмы действия продуктов вирусной активации . Примером такого продукта является протеинкиназа, активируемая геном sre, а также целый ряд других вирус-индуцированных канцерогенов. Это и рецепторы к эпидермальному фактору роста, продуцируемые геном v-erb, и тромбоцитарный фактор роста (ТФР), кодируемый фрагментами гена v-sis, и целый ряд связывающихся с ядром клетки белков, продукцию которых вызывает вирус птичьей лейкемии.
Вирусные онкогенные молекулы в большинстве случаев структурно отличаются от своих аналогов клеточного происхождения; кроме того, в них отсутствуют интроны. Например, белок, кодируемый геном v-erb, гомологичен клеточному рецептору к эпидермальному фактору роста (ЭФР), но в нем отсутствует часть экстраклеточного домена, включая ЭФК-специфичный сайт. Так как в продуцируемой вирусом молекуле отсутствует плазматический участок, ответственный за автофосфорилирование, такой вирусный рецептор все время находится во «включенном» состоянии.
Как нормальные, так и раковые клетки содержат в себе участки последовательностей ДНК, гомологичные РНК онкогенных вирусов. Если происходит экспрессия или активация под действием канцерогенов этих клеточных онкогенов, это ведет к злокачественной трансформации клеток.
 Онкогенная и злокачественная трансформация.
Онкогенная и злокачественная трансформация.
На стадии А нормальная клетка, которая характеризуется низкой протоонкогенной активностью, продуцирует фактор роста (х) или белки дифференцировки или рецепторы (у).
Канцерогены повышают активность протоонкогенов, что дает начало неопластической трансформации.
По другому механизму: при заражении ретровирусом происходит внедрение в ДНК клетки вирусных промоторов или онкогенов (Б), что также ведет к увеличению онкогенной активности и последующей злокачественной трансформации.
Вирус может активировать процессинг в клетках путем внедрения в их специальных регуляторных последовательностей - промоторов считывания белка, нарушая таким образом нормальные процессы транскрипции. Запускаемый таким образом механизм внедренного мутагенеза может вовлекать в себя целый комплекс различных процессов. Примером может служить внедрение в ДНК клетки вирусной после довательности «многочисленных терминальных повторений» (МТП). При внедрении в клеточную ДНК данная последовательность инициирует транскрипцию в обоих направлениях цепочки ДНК, позволяя транскрибировать одновременно как клеточные, так и вирусные гены. Такой механизм действия характерен для вируса клеточного лейкоза, когда вирусная ДНК интегрируется в клеточную сразу вслед за участком с-тус, вызывая его активацию.
Первым ретровирусом , для которого была однозначно доказана связь со злокачественными новообразованиями, был вирус Т-клеточной лейкемии человека (ВТКЛ-1), выделенный из клеток хронической кожной Т-лимфомы. Этот вирус довольно широко распространен, может передаваться половым путем, через кровь, особенно в среде наркоманов, и от беременной женщины к плоду. Первоначально эндемический ареал циркуляции данного вируса был в основном представлен тропическими странами, однако в настоящее время в США серопозитивная реакция на вирус обнаруживается у каждого из 4000 человек населения. Кроме Т-клеточной лейкемии вирус вызывает тропический спастический паралич.
После 20 лет наблюдения за серопозитивными пациентами оценено, что риск развития последнего заболевания составляет около 5%. Один из вирусных генов, а именно tax-ген, вызывает увеличение продукции клеточного интерлейкина-2 (ИЛ-2) и его рецепторов, что является основным фактором, стимулирующим деление Т-клеток.
Ретровирусы могут вызывать опухолевые заболевания не напрямую, а опосредованно, как это было показано для вируса иммунодефицита человека (ВИЧ-1), вызывающего развитие СПИДа. Случаи развития рака у ВИЧ-инфицированных рассмотрены в одном из обзоров. Отмечается, что у ВИЧ-инфицированных наиболее часто развиваются три типа опухолей: мгновенная или высокозлокачественная В-клеточная лимфома; саркома Капоши (СК, которую вызывает другой вирус - герпесвирус ГСК, или герпесвирус 8); карцинома шейки матки.
До разработки эффективных методов терапии ВИЧ-инфицированных более 40% из них заболевали тем или иным видом рака. Тем не менее связь этого вируса с развитием онкологических заболеваний скорее всего косвенная и опосредована развитием общей хронической иммунодепрессии организма, которая и позволяет другим канцерогенным вирусам вызывать рак. В-клеточные лимфомы также характеризуются довольно сложным патогенезом. Хотя В-клетки не поражаются вирусом ВИЧ-1, они могут быть мишенью для других типов вирусов, например для вируса Эпштейна-Барра (ВЭБ). Рак шейки матки у женщин также развивается на основе вторичной вирусной инфекции - вируса папилломы человека (ВПЧ), на фоне общей иммунодепрессии, вызванной ВИЧ-1. По причине общего снижения иммунных реакций организма все эти опухоли развиваются особенно быстро и агрессивно.
Похожие статьи